Yuan-Qi He1,2,
Hai-Ning Liu2,
Xiao-Xiao Li2,
Guo-Yun Wang3 ![]()
For correspondence:- Guo-Yun Wang Email: doctorguoyun@126.com Tel:+8653182169114
Received: 4 January 2016 Accepted: 17 May 2016 Published: 28 June 2016
Citation: He Y, Liu H, Li X, Wang G. Antitumor activity of physcion 8-o-^6;-glucopyranoside against cervical cancer by induction of apoptosis. Trop J Pharm Res 2016; 15(6):1145-1150 doi: 10.4314/tjpr.v15i6.5
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To investigate the antitumor activity of physcion8-O-β-glucopyranoside (PSG) against cervical cancer, as well as its mechanisms.
Methods: The anti-proliferative effects of PSG on HeLa cells were determined by CCK-8 assay and the half maximal inhibitory concentration (IC50) values were calculated. Subsequently, a mouse xenograft model of HeLa cell line was established to investigate the antitumor effect of PSG in vivo. Furthermore, cell apoptosis was investigated by @258;uorescence microscopy via DAPI staining, and other mechanisms were determined by Western blot assay.
Results: In vitro, PSG exhibited significant anti-proliferative effect on HeLa cells (p <0.05) in concentration-and time-dependent manners, with an IC50 value of 41.34 μg/mL. In vivo, PSG also had significant anti-tumor activity in nude mouse xenograft model (p < 0.05), inhibiting tumor growth. Furthermore, the results showed that treatment with PSG (20, 40 and 60 μg/mL) for 24 h resulted in significantly increased apoptosis in HeLa cells (p < 0.05). Additionally, Western blot analysis revealed that after exposure to 20, 40 and 60 μg/mL of PSG for 24 h, protein ex
Conclusion: The results demonstrate that PSG possesses notable anti-tumor activity against cervical cancer and that the mechanisms involve induction of apoptosis by mitochondria-mediated signaling pathway.
Introduction
In spite of the improvements that have been achieved in diagnostic and screening techniques, cervical cancer still remains the fourth leading cause of cancer related deaths among women worldwide and the second cause of deaths among women in developing countries [1,2]. The average age of patients with recurrent cervical carcinoma is about 40 - 45 years [3]. In addition, the recurrence for stages III and IVA is 42 and 74 %, respectively. At present, chemotherapy or neoadjuvant chemotherapy is always the first choice for those patients with advanced cervical cancer, whereas the overall survival remains poor [4]. Therefore, there is an urgent need to find effective drugs or therapeutic methods to treat cervical cancer. It has been reported that natural products derived from plants and microorganisms have anticancer activities, which provide useful clues for discovering new anticancer drugs.
Physcion 8-O-β-glucopyranoside (PSG) is an anthraquinone () extracted from Rumex japonicas Houtt [5]. Rumex japonicas Houtt, a perennial herb plant belonging to the family Polygonaceae, is widely distributed in China and recorded [6]. It has been traditionally used in folk medicine for relieving bowel movement, stopping bleeding and as an antipruritic agent [6]. At present, it is recognized that R. japonicas has new pharmacological activities such as anti-microorganic, anti-oxidant, anti-inflammatory and anti-tumor [7-10]. In addition, a large number of bioactive compounds have been found in R. japonicas, including anthraquinones, emodin, chrysophanol, physcion, fatty acids, condensed tannins, and so on [11-13].
PSG is one of the major bioactive compounds present in R. japonicas, and has been reported to have significant anti-proliferative activity on A549 cell lines by inducing apoptosis and cell cycle arrest [5]. However, to the best of our knowledge, the anti-cervical cancer effects of PSG isolated from R. japonicas has not been researched at present. In our present work, we focus on the anti-tumor effect of PSG in human cervical cancer cell line HeLa in vitro and in vivo, and explore the possible mechanisms, which may provide a scientific basis for treating this disease in clinic.
Methods
Chemicals
Physcion 8-O-β-glucopyranoside (PSG) was purchased from the Sichuan Keluoma Biotech. Co. (Chengdu, China) and was isolated from the dry root of R. japonicas with purity of more than 98 %. Dulbecco's Modified Eagle Medium (DMEM) and fetal bovine serum (FBS) were purchased from Life Technologies Inc (Gibco BRL); Penicillin-Streptomycin Solution (100X), cell counting kit-8, cell lysis buffer for Western and IP, BCA protein assay kit, BeyoECL Plus, 2-(4-Amidinophenyl)-6-indolecarbamidine dihydrochloride (DAPI), horseradish-peroxidase (HRP) -labeled goat anti-rabbit IgG (H+L) were purchased from Beyotime (Haimen, China); cleaved-caspase-3 (c-caspase-3), cleaved-caspase-9 (c-caspase-9) antibodies were purchased from Cell Signaling Technology (Beverly, MA, USA); Bcl-2, Bax and β-actin antibodies were purchased from Abcam Biotechnology (Cambridge, MA, USA); all other chemicals used in this study were of analytical grade.
Cell culture
Immortalized human cervical cells (HeLa cells) were obtained from the Shanghai cell bank of Chinese academy of sciences (Shanghai, China). Cells were cultured in DMEM medium supplemented with 10 % FBS and antibiotics (100 U/mL penicillin G and 100 mg/mL of streptomycin) and maintained in a humidified atmosphere containing 5 % CO2 at 37 ºC.
Cell viability assay by CCK-8
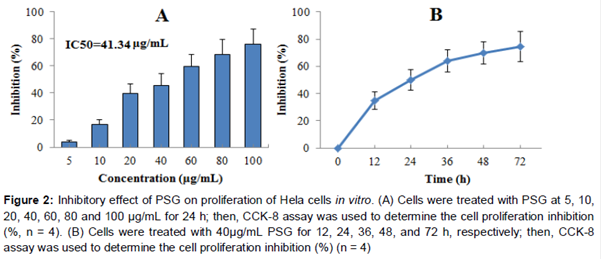
Cell viability was carried out using the CCK-8 assay. In Brief, HeLa cells were cultured until they reached approximately 90 % confluency before preparing the plates for the cell viability assay. Cells were seeded in 96-well plates at 5 × 103 cells per well and grown for 24 h. Then, cells were treated with various concentrations of PSG (5, 10, 20, 40, 60, 80 and 100 μg/mL) for 24 h and then the cell proliferation inhibition was determined by CCK-8 following the manufacturer’s instructions. Optical density values (OD) were measured at 570 nm by a microplate reader (Bio-Rad, CA, USA) and the IC50 value of PSG on HeLa cells was calculated. Furthermore, for determining the time-dependent manner, Hela cells were treated with PSG (40 μg/mL) for 12, 24, 36, 48, and 72 h, respectively; then, CCK-8 assay was used to determine the cell proliferation inhibition. The inhibition rate was calculated according to Eq 1.
Inhibition (%) = {(Ac – At)/Ac}100 …………. (1)
where Ac and At are the absorbance of control and treated samples, respectively.
Xenograft model of nude mice
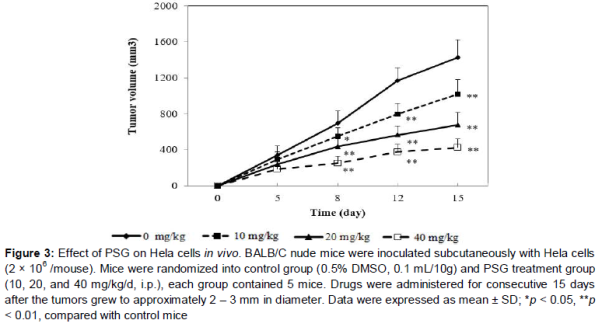
The antitumor effect of PSG in vivo was evaluated with the animal tumor model by inoculation of HeLa cells as previously reported [14]. BALB/C nude mice were randomly divided into control group (0.5 % DMSO, 0.1 mL/10g) and PSG treatment group (10, 20, and 40 mg/kg/d, i.p.), each group contained 5 mice. Nude mice (18-22 g) were injected subcutaneously in the right flank with HeLa cells (2.0 × 106/mouse).
When the tumor grew to approximately 2 – 3 mm in diameter, mice were treated with various concentrations of PSG (intraperitoneal injection, i.p.) and equal volume of solvent control (0.5 % DMSO, i.p.), respectively. Drugs were treated for consecutive 15 days and tumor sizes (width and length) were measured at 0, 5, 8, 12 and 15 days. Finally, the tumor volumes were determined using the formula: volume = (width2 × length)/2.
Nuclear staining with DAPI
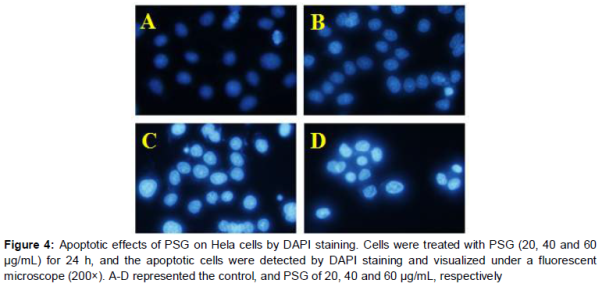
Cell apoptosis analysis was determined by DAPI staining assay. About 1 × 105 HeLa cells per well were cultured in 12-well plates and were treated with PSG (20, 40 and 60 μg/mL) for 24 h. Then, cells were stained with 10 μg/mL of DAPI for 10 min at 37 °C, in the dark. Finally, the changes in the cells’ nucli were observed with a fluorescence microscope (Olympus, IX53, Tokyo, Japan).
Western blot assay
HeLa cells (1 x 106 cells/flask) were seeded in T-25 flasks and were treated with PSG 20, 40 and 60 μg/mL of PSG, then 24 h later, cells were harvested and total proteins were extracted with cell lysis buffer for western and IP. Subsequently, the protein concentration was determined by the BCA protein assay kit. Then, equal amount of total protein (40 μg) were separated by the sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS/PAGE). After that, all proteins were transferred to PVDF membranes and probed withcaspase-3, caspase-9, Bax, Bcl-2 and β-actin monoclonal antibodies, respectively. After 12 h of incubation at 4 °C, HRP labeled secondary antibody was added and incubated for 2 h at 37 °C. Finally, protein bands were detected by Beyo ECL Plus kit (chemiluminescence). To normalize for protein loading, the Bio-Rad quantity one software was used to quantify the blot density and the proteins expression levels were expressed as a relative value to that of β-actin.
Statistical analysis
All values are represented as mean ± standard deviation (SD, n = 3). Statistical analysis was performed by two-tailed Student’s t-test using SPSS 19.0 software, and differences were considered significant at p < 0.05.
Results
Inhibitory effect of PSG on cervical cancer in vitro
The anti-cervical cancer activity of PSG was determined by CCK-8 in HeLa cells. As can be seen from A, PSG exhibited significant anti-proliferative effects in concentration-dependent manner on HeLa cell, and the IC50 value was 41.34 μg/mL. Furthermore, the anti-proliferative effect of PSG was also time-dependent (B).
Inhibitory effect of PSG on cervical cancer in vivo
To further explore the anti-cervical cancer activity of PSG in vivo, PSG treatment was performed in the nude mouse xenograft model with HeLa cells. As shown in , PSG (10, 20 and 40 mg/kg, ip) significantly reduced tumor volume as compared to the solvent control group (p < 0.05, p < 0.01, p < 0.01), especially at dose of 40 mg/kg and at day 15.
Pro-apoptotic effect of PSG on HeLa cell
According to the results above, PSG showed obvious anti-tumor activity on the human cervical cancer cell in vitro and in vivo. To explore whether the anti-tumor activity of PSG was related to apoptosis, HeLa cells were stained with DAPI. As shown in , after treatment with PSG (20, 40, 60 μg/mL), cells exhibited obvious apoptosis features such as improved brightness, nuclear condensation and nuclear fragmentation. This result suggested that the anti-proliferative effect of PSG might be related to induction of apoptosis.
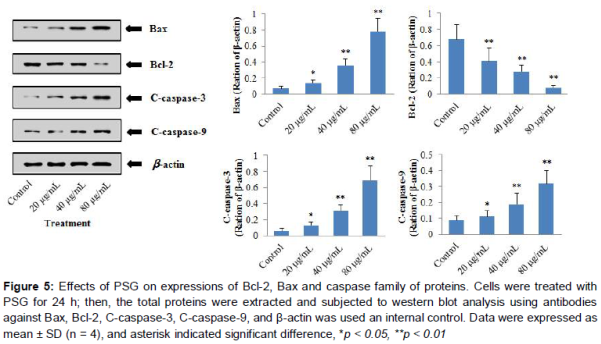
Effect of PSG on protein expression of caspase-3, caspase-9, Bax, Bcl-2 in HeLa cell
To further investigate the possible mechanism by which PSG induces apoptosis in HeLa cells,C-caspase-3, C-caspase-9, Bcl-2 and Bax protein expression levels were studied by western blot. As shown in , there was significant difference between untreated HeLa cells and PSG treatment group for all these protein expressions. Interestingly, compared to the control group, the protein expression levels of Caspase-3, Caspase-9 and Bax were gradually up-regulated with increased concentrations of PSG (20, 40, 60 μg/mL, p < 0.05, p < 0.01, p < 0.01, respectively), whereas Bcl-2 protein expression was down-regulated significantly (p < 0.01) in a concentration-dependent manner.
Discussion
To the best of our knowledge, this study is the first report regarding the antitumor effect of PSG on human cervical cancer cell line in vitro and in vivo. Furthermore, we first investigated the possible mechanisms of PSG on human cervical cancer cell line HeLa in our present study. The findings of the present study showed that PSG significantly inhibited cell proliferation in concentration-and time-dependent manners in HeLa cells. And PSG also reduced the tumor growth in nude mouse xenograft model with HeLa cells in a dose-dependent manner. In addition, we also found that exposure of HeLa cells to PSG leads to a marked increase in apoptosis, indicating that apoptosis contributes to growth inhibition in PSG-treated HeLa cells.
Apoptosis, known as programmed cell death (PCD), is important for both health and disease and is one of the major mechanisms of cell death in response to cancer therapies [5,15]. Apoptosis mechanisms are highly complex. It is recognized that apoptosis is mediated by the extrinsic (death receptor) pathway, intrinsic (mitochondrial) pathway, and perforin/granzyme pathway [15].
Each pathway is initiated by its own signalling molecules like caspases. For example, the extrinsic, intrinsic and granzyme B pathways can be initiated by caspase-8, caspase-9 and casepase-10, respectively. Once caspases are activated, the apoptotic signaling pathway will be activated resulting in cell death. Caspase-3 is the most important effector or executioner of the extrinsic, intrinsic and granzyme B pathway [15,16]. In our experiment, PSG significantly increased the protein expression of cleaved-caspase-3 and cleaved-caspase-9 in HeLa cells, which indicated that the PSG-induced apoptosis process is due to activation of the mitochondrial pathway.
Furthermore, it has been reported that the Bcl-2 family of proteins play a critical role in control and regulation of mitochondrial mediated apoptotic pathway, because the mitochondrial membrane permeability is governed by this family of proteins [17]. The Bcl-2 family proteins have pro-apoptotic and anti-apoptotic effects. Bax is a critical pro-apoptotic protein while Bcl-2 is an important anti-apoptotic protein [18]. Bcl-2 is an integral membrane protein, which causes anti-apoptosis mainly by preventing mitochondrial outer membrane permeabilization. However, if the effect of Bcl-2 is suppressed, Baxoligomerization takes place and thus leads to apoptosis. Activation of Bax can result in the release of the contents of the mitochondrial inter-membrane space, which then activates caspases and leads to a series of cascade reactions that result in cell death [17]. Our results demonstrated that exposure of PSG to HeLa cells leads to up-regulation of Bax protein while it down regulates Bcl-2 protein.
Conclusion
The findings of the present study suggest that PSG possesses antitumor effect against human cervical cancer cell line by inducing mitochondrial-mediated apoptosis via up-regulation of caspase-3, caspase-9 and Bax, as well as down-regulation of Bcl-2.
References
Archives


News Updates